INTRODUCTION
Plants, just like other photosynthetic organisms, are capable of capturing the light energy and using it to produce their food by photosynthesis. This may sound simple but in fact it is a very complex process. However, the process of converting light energy can only be performed in the green parts of the plants like leaves. The reason behind this is that not all cells of plants have chloroplast, which is the site of photosynthesis. The non-green parts like roots and stem have other forms of plastids, modified either to store molecules (leucoplast) or to provide color (chromoplast).
The ground tissue of leaves called Mesophyll is the main site of photosynthesis in plants. Each of these cells contain as many as 30-40 chloroplast capable of producing food. A chloroplast is a cell orgenelle of variable shapes and sizes surrounded by two membranes externally. The outer membrane and the internal membrane are separated by a small space called Intermembrane space. The fluid part enclosed inside the internal membrane is called Stroma which contains several contents like the circular double stranded DNA of Chloroplast which makes it semi-autonomous, all major types of RNA, ribosomes (70S type), and many proteins and enzymes. Along with it the stroma also contains membrane bound sac like structures called Thylakoids. In higher plants, the thylakoids are stacked over each other to form Grana and the granal thylakoids are connected through membranous extensions called Stroma lamellae.
GRANA AND STROMA LAMELLAE
PHOTOSYSTEMS
The light capturing pigments are present in the thylakoid and stroma lamellae in the form of groups called Photosystems. Each photosystem is a group of 200-300 pigments called accessory pigments (forming the Light Harvesting Complex) and also includes a special type of Chlorophyll a (dimeric) as its Reaction center. On the basis of type of reaction center, the photosystems are divided into two types- Photosystem I and Photosystem II. The names I and II are given on the order of their discovery.
1) PHOTOSYSTEM I
- The photosystem I is present on the non-appressed parts of the grana as well as the stroma lamellae.
- The ratio of chlorophyll a to chlorophyll b is high in the LHC of photosystem I
- The reaction center chlorophyll is designated as P700 as its peak absorption occurs at 700nm wavelength of light.
2) PHOTOSYSTEM II
- The photosystem II is present on the appressed parts of the grana only.
- The ratio of chlorophyll a to chlorophyll b is almost equal in the LHC of photosystem II.
- The reaction center here is called P680 due to its peak absorption at 680nm wavelength.
- The photosystem II is also associated with a water splitting complex or also called Oxygen Evolving Complex(towards the thylakoid lumen) capable of splitting water molecules into protons, electrons and oxygen. The OEC uses Ca ions, Mn ions and Cl ions as cofactor and releases one electron at a time to the P680.
CYTOCHROME COMPLEX
Besides the photosystem, the thylakoid and stroma lamellae also contains a cytochrome complex called cytochrome b6f complex (similar to complex III or the mitochondrial ETS). This complex is equally distributed in both appressed and non-appressed parts of grana. The cytochrome b6 f complex contains a b-type cytochrome with two heme groups (designated bH and bL), a Rieske iron-sulfur protein and cytochrome f.
ELECTRON CARRIERS
Several electron carriers are also present in the membranes for helping in the flow of electrons from one photosystem to the other. These electron carriers are either of these two types-
1) PLASTOQUINONE
Plastoquinone or PQ is a lipid soluble electron carrier (can freely diffuse in lipid bilayer) which receives electrons coming from Photosystem II and transfers it to cytochrome b6f complex. It is also responsible for pumping protons from stroma side to the lumen thus creating a proton gradient required for ATP formation.
2) PLASTOCYANIN
Plastocyanin or PC is another electron carrier of the thylakoid membrane. It is a copper containing water soluble electron carrier present towards the lumen side of the membrane. The plastocyanin receives electrons from cytochrome b6f complex and transfers it to photosystem I.
ENZYMES
1) ATP SYNTHASE
The thylakoid and stroma lamellae also contains an enzyme having ATP synthase activity for Photophosphorylation. The activity of this ATP synthase is similar to that of one found in the inner mitochondrial membrane. It also consist of two parts- CF1 and CF0 where CF1 has main catalytic activity and undergoes conformational changes to make ATP while the CF0 simple acts as an ion channel for facilitated diffusion of protons. The CF1 part is located towards the stroma side.
2) FERREDOXIN:NADP+ OXIDOREDUCTASE
The Ferrodoxin: NADP+ oxidoreductase is a flavoprotein present on the thylakoid membrane and it reduces the NADP+ to NADPH by capturing electrons from reduced ferredoxin and transferring it to NADP+.
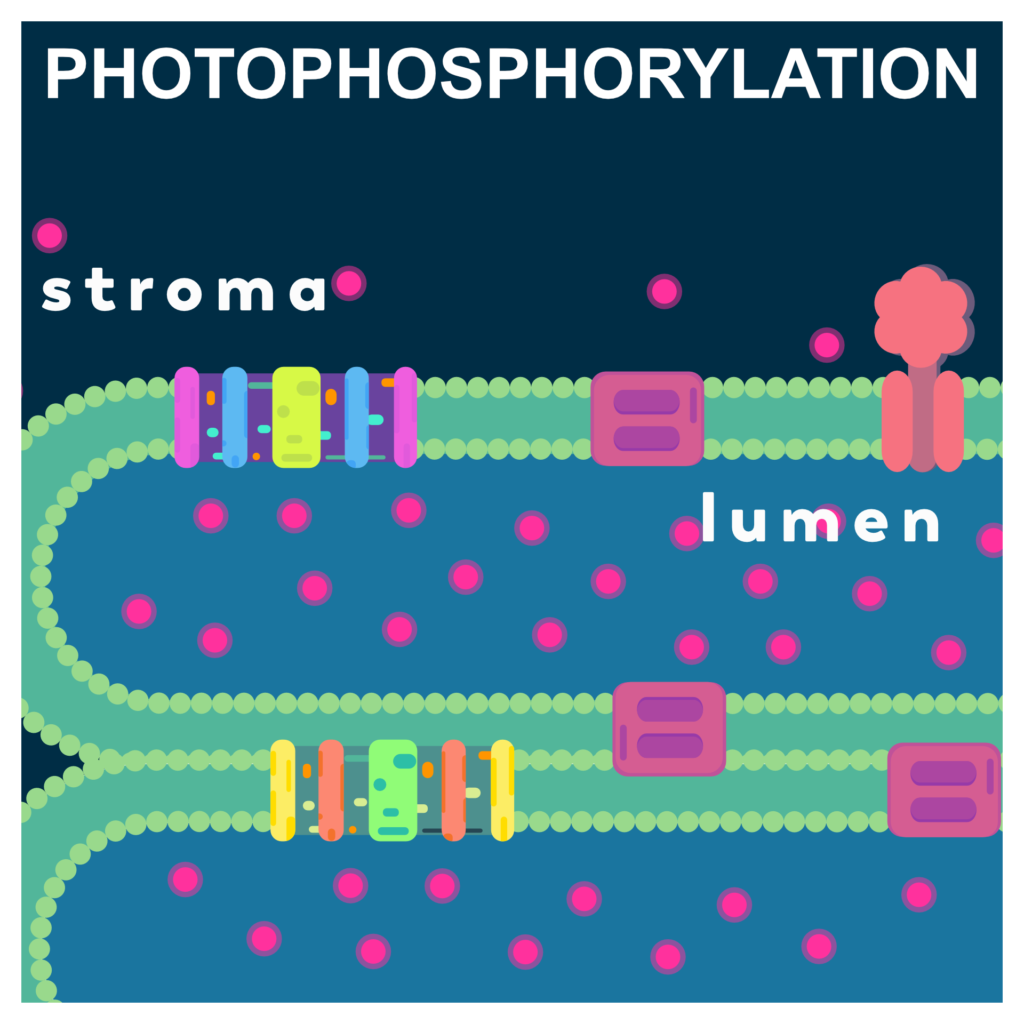
ELECTRON TRANSPORT AND PHOTOPHOSPHORYLATION
Photophosphorylation is the process of formation of ATP by using light energy. Just like oxidative phosphorylation , photophosphorylation also depends on proton gradient to produce ATP form ADP and inorganic phosphate.
The process starts as soon as the light strikes the leaves. The light harvesting complex of each photosystem absorbs lights of different wavelengths and transfers this energy to the reaction center of the respective photosystem. As a result, the reaction center acquires sufficient light energy, its electrons reaches an excited state and is captured by the respective associated electron carriers.
The photoexcitation of electrons from Photosystem II reaction center is captured by the nearest Plastoquinone which is tightly bound called PQ-a and then to a loosely bound PQ-b. The PQ-b also acquires protons from the stroma side and get reduced to plastoquinol (PQ-bH2). The loss of electron from PS II is compensated by splitting water molecules in the lumen by the OEC.
The electrons are then transferred from PQbH2 to cytochrome b6f complex and the protons are released inside the lumen. The cytochrome complex receives the electrons and passes it to cyt b6-FeS-cyt f chain and then these electrons are captured by the plastocyanin (PC) located towards lumen side of the membrane.
Photoexcitation of electrons also occurs from PS I creating a deficiency in PS I reaction center. The electrons from PC balances this deficiency by releasing the electrons captured from cytochrome b6f complex. Electrons released from PS I reaction center are transferred to an iron sulfur protein and then to Ferredoxin (Fd). These electrons from Fd (reduced) are then captured by an enzyme ferredoxin: NADP+ oxidoreductase located towards the stroma side of the thylakoid membrane. This enzyme transfer electrons from Fd (reduced) to NADP+ thus reducing it to NADPH.
The proton gradient created by the flow of electrons from PS II to PS I creates a p-side in lumen and a n-side in stroma. This proton gradient breaks by facilitated diffusion of protons from lumen to stroma through CF0 of the ATP synthase complex. The CF1 part of ATP synthase undergoes conformational changes and phosphorylates ADP to form ATP.